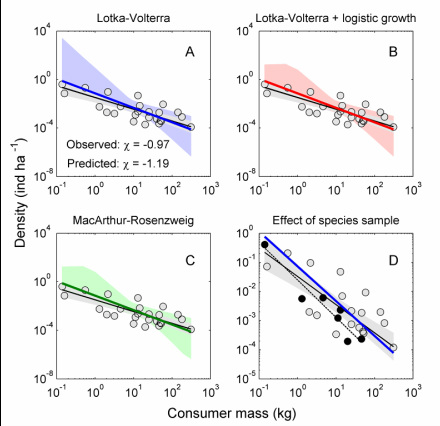
The abundance of organisms tends to decrease with body size. Mice are common; elephants not so much. On the one hand it seems obvious that large organisms need more space and food, so there might be fewer of them. But it isn't necessarily so, as it depends on the resources available to organisms and how they use them (for example, larger organisms could be better at accessing food then smaller organisms). So how can we explain and perhaps predict the slope of the typically negative relationship between body size and abundance?
We took a simple approach to this problem. By making the parameters of a simple predator-prey model 'scale' with body size, we could solve for the steady-state density as an explicit function of size. We then collected data on how model parameters (e.g., Lotka-Volterra type parameters) changed with size, plugged that information into our model, and compared the model prediction with real data on the density of organisms. We focused on two groups: terrestrial mammalian carnivores and aquatic grazing protists. For both groups, the simplest possible predator-prey model (Lotka-Volterra) gave a near-perfect prediction of observed relationships between size and density for these groups. We are using this approach to investigate the abundance scaling in other groups and extending the approach to other types of problems such as trophic cascades.
See:
DeLong, J.P., and ten others. 2015. The body size dependence of trophic cascades. American Naturalist 185:354-366.
DeLong, J.P. and D.A. Vasseur. 2012. Size-density scaling in protists and the links among consumer-resource interaction parameters. Journal of Animal Ecology. 81:1193-1201.
DeLong, J.P. and D.A. Vasseur. 2012. A dynamic explanation of size-density scaling in carnivores. Ecology. 93:470-476.
We took a simple approach to this problem. By making the parameters of a simple predator-prey model 'scale' with body size, we could solve for the steady-state density as an explicit function of size. We then collected data on how model parameters (e.g., Lotka-Volterra type parameters) changed with size, plugged that information into our model, and compared the model prediction with real data on the density of organisms. We focused on two groups: terrestrial mammalian carnivores and aquatic grazing protists. For both groups, the simplest possible predator-prey model (Lotka-Volterra) gave a near-perfect prediction of observed relationships between size and density for these groups. We are using this approach to investigate the abundance scaling in other groups and extending the approach to other types of problems such as trophic cascades.
See:
DeLong, J.P., and ten others. 2015. The body size dependence of trophic cascades. American Naturalist 185:354-366.
DeLong, J.P. and D.A. Vasseur. 2012. Size-density scaling in protists and the links among consumer-resource interaction parameters. Journal of Animal Ecology. 81:1193-1201.
DeLong, J.P. and D.A. Vasseur. 2012. A dynamic explanation of size-density scaling in carnivores. Ecology. 93:470-476.
When predators move through space, they encounter not just prey but other predators. This process leads to interference competition. Because of the way that movements are altered by the presence of other predators, there is a nonlinear (and quite powerful) affect of increasing predator density on foraging rates. Interference is one of those profoundly important processes that is mostly ignored because considering it takes a fair bit more work. For example, if you want to measure the functional response of a particular predator-prey pair, you have to triple or quadruple your work load to also measure foraging at different levels of predator density. So, for the thousands of functional responses that have been measured, there are only a few dozen or so that have explicitly included interference competition. Nonetheless, whenever people look for interference, they invariably find it. In my lab, we try to measure interference when possible (remaining understanding about times when it is simply too challenging) and investigate the causes and consequences of its magnitude.
See:
DeLong, J.P., T.C. Hanley, and D.A. Vasseur. 2014. Predator-prey dynamics and the plasticity of predator body size. Functional Ecology. 24:487-493.
DeLong, J.P. and D.A. Vasseur. In press. Linked exploitation and interference competition drives the variable behavior of a classic predator-prey system. Oikos.
DeLong, J.P., T.H. Hanley, and D.A. Vasseur. 2013. Competition and the density dependence of metabolic rates. Journal of Animal Ecology.
DeLong, J.P. and D.A. Vasseur. 2011. Mutual interference is common and mostly intermediate in magnitude. BMC Ecology 11:1.
See:
DeLong, J.P., T.C. Hanley, and D.A. Vasseur. 2014. Predator-prey dynamics and the plasticity of predator body size. Functional Ecology. 24:487-493.
DeLong, J.P. and D.A. Vasseur. In press. Linked exploitation and interference competition drives the variable behavior of a classic predator-prey system. Oikos.
DeLong, J.P., T.H. Hanley, and D.A. Vasseur. 2013. Competition and the density dependence of metabolic rates. Journal of Animal Ecology.
DeLong, J.P. and D.A. Vasseur. 2011. Mutual interference is common and mostly intermediate in magnitude. BMC Ecology 11:1.

This is my exciting new predator-prey system. Actinosphaerium catches Paramecium and other protists with long sticky spikes, which are then pulled back into the cell with the prey attached. Each prey gets its own vacuole. One great thing about this system is that you can see the prey inside the cells if the prey are a dark color (see green vacuoles in cells, each with a single Paramecium inside), allowing one to quantify predation by counting vacuoles. It also is handy because the predators do not move much and are big enough to count easily in flat microcosms. The Actinosphaerium are highly plastic in size, meaning that the system is ideal for linking predator size to foraging behavior and population dynamics and temperature. Experiments in these arenas are ongoing.
See:
DeLong, J.P. 2012. Experimental demonstration of a rate-size trade-off governing body size optimization. Evolutionary Ecology Research. 14:343-352.
See:
DeLong, J.P. 2012. Experimental demonstration of a rate-size trade-off governing body size optimization. Evolutionary Ecology Research. 14:343-352.
